 Darwin of the gaps
Darwin of the gaps
- (1) fra God and Evolution; J.Wells
I juni 2000 annonserte president Bill Clinton at den første inspeksjonen av det menneskelige genom, DNA-sekvensen til en menneskelig celle, var fullført. Han uttalte: "i dag lærer vi språket Gud brukte da han skapte livet." Ved presidentens side var Fr. Collins, leder av Human Genome Project, som hadde hjulpet å skrive Clintons tale. "Det er ydmykende og i dyp respekt" sa han, "å innse at vi har fått det første innsyn i vår instruksjonsbok, tidligere bare kjent av Gud." Den komplette gen-sekvensen ble annonsert tre år senere, og i 2006 publiserte Collins en bok: "The language of God": A Scientist Presents Evidence for Belief.
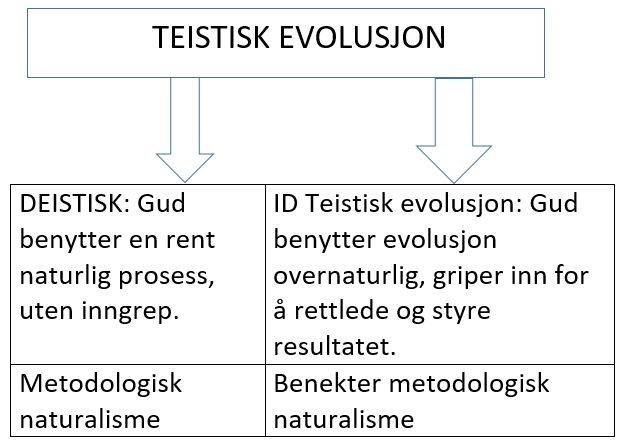
Bilde 1. (Minst) to typer av teistisk evolusjon
Ironisk nok, inneholdt ikke Collins bevis DNA. Det mente han forsynte tvingende bevis for Darwins evolusjonsteori i stedet. Han uttalte at Darwins evolusjonsteori "kunne og måtte være sann" og at folk som argumenterer for at den feiler i å forklare det åpenbare designet i levende organismer, klynger seg til en 'Gud i hullene' teori. Han uttalte at den eneste vitenskapelige overensstemmede og åndelig tilfredsstillende måte å være en troende på, er via Teistisk evolusjon eller BioLogos.
Likevel ignorerer Collins bevis, som han kom til å gjøre flere ganger, ut fra prosjektet han ledet. I stedet for å støtte Darwinistisk evolusjon (DE), underminerer det nye DNA-beviset den. Dess mer vi lærer om vårt genom, dess mindre holdbar blir DE.
Gud i hullene
I sin bok "The language of God" baserer Collins primært sin kristne tro på den moralske lov, forestillingen om rett og galt, som alle mennesker innehar. Han ser også grunner for tro i den orden naturlovene framstiller, det antropiske prinsipp og i Big Bang. Faktumet at universet hadde en begynnelse, gjør at det må sitt opphav utenfor seg selv. Universets fininnstilling, en serie av bemerkelsesverdige sammentreff, som tillater naturen å støtte liv. Men Collins advarer mot en 'Gud i hullene' teori. Tro som plasserer Gud i hullene av vitenskapelig forståelse, kan gå mot krise om vitenskapelig framgang fyller hullene.
 En skeptiker kan innvende at Collins selv synes å fylle hullene med sine forestillinger (sml. Confirmation Bias/Bekreftelsestendens). Han skriver: "DNA alene, kan aldri forklare noe slikt som den moralske lov, selv om den ble fulgt opp av en svær ansamling vitenskapelige data om DNA's funksjonalitet. Og Big Bang roper ut om en guddommelig inngripen. Jeg kan ikke se hvordan naturen kan ha skapt seg selv." Selvsagt tror noen evolusjons-psykologer at de allerede har forklaringen på den moralske lov, som et produkt av Darwinske og Freudske prosesser. Og noen fysikere tror de har forklart Big Bang som et resultat av 'cykliske ekspansjoner og kontraksjoner i et evig univers', eller som begynnelsen på 'ett av talløse andre multivers'.
En skeptiker kan innvende at Collins selv synes å fylle hullene med sine forestillinger (sml. Confirmation Bias/Bekreftelsestendens). Han skriver: "DNA alene, kan aldri forklare noe slikt som den moralske lov, selv om den ble fulgt opp av en svær ansamling vitenskapelige data om DNA's funksjonalitet. Og Big Bang roper ut om en guddommelig inngripen. Jeg kan ikke se hvordan naturen kan ha skapt seg selv." Selvsagt tror noen evolusjons-psykologer at de allerede har forklaringen på den moralske lov, som et produkt av Darwinske og Freudske prosesser. Og noen fysikere tror de har forklart Big Bang som et resultat av 'cykliske ekspansjoner og kontraksjoner i et evig univers', eller som begynnelsen på 'ett av talløse andre multivers'.
Collins griper ikke fatt i slike 'forklaringer', men griper fatt i hva han synes er den mest ekstreme 'Gud i hullene' teori: Intelligent Design (ID) i biologien. Collins stiller opp en 'stråmann-forestilling' av ID, som han så angriper: ID hviler på tre forestillinger: i) evolusjon fremmes som et ateistisk verdenssyn ii) evolusjon viser svære mangler når det ikke kan gjøre rede for intrikat kompleksitet i naturen. iii) om ikke evolusjon kan forklare ikke-reduserbar-kompleksitet, må det være intelligent design (ID). Men ingen av disse tre representerer presist hva ID står for. I følge Discovery instituttets Senter for Vitenskap og Kultur, så holder ID for at visse trekk ved universet og levende organismer best kan forklares ved en intelligent årsak, ikke som en ikke-styrt prosess av naturlig utvalg.
Bilde 2. Beste forklaring
For å ta for oss Collins tre punkter, så holder ikke ID for at 'evolusjon fremmer et ateistisk verdenssyn'. Evolusjon kan bety noe så enkelt som endring over tid, som ikke innebærer noen implikasjoner i forhold til design. Evolusjon kan også bety mindre endringer innen eksisterende arter, noe som stadig observeres omkring i verden. Det er ikke generelt evolusjon, men Darwinismens ekskludering av design, som ID går mot. Darwin skrev at han ikke så mer design i variasjonen innenfor levende organismer eller i måten naturlig utvalg virker, enn i retningen vinden blåser. {Meteorologer ville her tilføye at vindretning går fra Høytrykks-områder mot lavtrykks-områder og bestemmes i tillegg av jordrotasjon m.m. Således er det klare begrensninger m.h.t. vindretning, hvorvidt den er tilfeldig slik Darwin syntes å forstå det -oversetters tilføyelse}. Vi skal la det ligge, og i stedet se på Collins andre og tredje utsagn:
Disse indikerer feilaktig at ID bare inneholder negative argumenter. At Darwinsk teori mangler forklaringsevne er et nødvendig, men ikke tilstrekkelig argument for å hevde ID. ID henfører til design bare når effektene ligner de som vi av erfaring kjenner til, krever en intelligent årsak. Som eks. på slikt kan nevnes M. Behes ikke-reduserbar-kompleksitet. Han beskrev det i 1996 som "et enkelt system, satt sammen av atskillige velkomponerte samvirkende deler, som samvirker til en basisfunksjon. Fjerning bare av én av komponentene vil føre til at systemet slutter å fungere." (5) Når vi møter slike systemer i hverdagen, slutter vi ganske fornuftig at de må skyldes en intelligent årsak/agent, som eneste årsak vi kjenner til å danne dem.
Behe beskrev flere slike ikke-reduserbart-komplekse systemer: blodets koaguleringsevne, et system av interagerende proteiner som sikrer at blod koagulerer bare når og hvor det er nødvendig. Men Collins forsikrer at 'det veletablerte fenomen ved gen-duplisering viser at komponent-delene til blod-koagulering, reflekterte gammel gen-duplisering som gradvis tillot den nye kopien å utvikle seg til å ta på seg denne nye funksjonen, drevet av naturlig utvalg. Behe skrev et bestemt svar på denne kritikken i 2000, men Collins ignorerte det.
Det mest ødeleggende for ID, i følge Collins syn, m.h.t. ikke-reduserbar kompleksitet er at nyere forskning angivelig skal underminere Behes argument i forhold til bakterie-flagellen. Den innehar et framdrifts system drevet av en mikroskopisk framdrifts-motor. Først omformulerer Collins Behes syn til at de individuelle delene til bakterie-flagellen ikke kunne hatt noen brukbar funksjon av noe slag. Så trekker Collins fram TTSS (et type III utsondrings-system). Det er et helt annerledes apparat, brukt av visse bakterier til å sprøyte gift inn i andre bakterier. TTSS er sammensatt av proteiner som ligner på noen av dem i bakterieflagellen. Collins skriver om denne: "Riktig nok er TTSS bare en del av flagellens puslespill, og vi vet ikke om vi noen gang kan fylle hele puslespillet, men hver slik ny bit i puslespillet gir en naturlig forklaring på noe ID hadde overlatt til overnaturlig årsaker. Det innebærer mindre og mindre territorium å stå for ID."
 Behe svarer for det første at "det ikke er noen grunn til at delene i ikke-reduserbare komplekse systemer ikke kan ha noen annen funksjon(6). Men igjen ignorerer Collins ham. Videre skrev Behe at bevis indikerer at bakterie-flagellen er eldre enn TTSS. Om det er noen sammenheng mellom dem, så må det være at TTSS 'devoluerte' fra bakterie-flagellen. Collins hovedargument mot ID er imidlertid 'gog's 'Gud i hullene' teori. Men den er ikke beskrivende for ID, men kanskje som begrunnelse for å forkaste animisme i naturen. Collins påstand at vitenskapelig framgang har benektet ID, er i det minste overdrevet. Alt han har kommet opp med er spekulasjoner om eldgammel gen-duplisering. Vi skal se på hvorfor ID ikke er en 'Gud i hullene' teori: ID trekker ikke inn noen overnaturlig intervensjon, om ikke effektene av intelligens i seg selv defineres som overnaturlig.
Behe svarer for det første at "det ikke er noen grunn til at delene i ikke-reduserbare komplekse systemer ikke kan ha noen annen funksjon(6). Men igjen ignorerer Collins ham. Videre skrev Behe at bevis indikerer at bakterie-flagellen er eldre enn TTSS. Om det er noen sammenheng mellom dem, så må det være at TTSS 'devoluerte' fra bakterie-flagellen. Collins hovedargument mot ID er imidlertid 'gog's 'Gud i hullene' teori. Men den er ikke beskrivende for ID, men kanskje som begrunnelse for å forkaste animisme i naturen. Collins påstand at vitenskapelig framgang har benektet ID, er i det minste overdrevet. Alt han har kommet opp med er spekulasjoner om eldgammel gen-duplisering. Vi skal se på hvorfor ID ikke er en 'Gud i hullene' teori: ID trekker ikke inn noen overnaturlig intervensjon, om ikke effektene av intelligens i seg selv defineres som overnaturlig.
ID påstår bare det minimalistiske krav at det er mulig å slutte fra evidens i naturen at noen mønstre eller egenskaper i naturen, kan forklares best ved en intelligente årsak, heller enn ikke-styrte prosesser. Så kan en selvsagt spørre hvordan designet er implementert, eller omkring naturen ved den intelligens det er snakk om. Til det siste ville det være naturlig å svare Gud, men ID tar oss ikke så langt. Den argumenterer ut fra vitenskapelige argumenter. Om en filosoferer over naturen ved den intelligens som står bak det hele, er en over i naturlig teologi.
Ellers er ikke design-argumenter ut fra uvitenhet. Ingen forstandig person argumenterer: 'Jeg vet ikke hva som forårsaket X, dermed må X være designet.' I dagliglivet slutter vi til design når X ligner noe vi vet er produsert intelligente prosesser og aktører. Ikke-reduserbar kompleksitet er ett kjennetegn ved designede artefakter, spesifisert kompleksitet (W.Dembski) er ett annet, eller om noe innehar preskriptiv informasjon (K.Tveter). I alle tilfeller, slutter vi mer pålitelig til design når vi har flere bevis på det enn om vi har færre.
Darwin og hans samtidige trodde levende celler inneholdt dråper av protoplasma. For dem var det lett å slutte at de ikke var designet, ut fra mangel på viten. Men i det biokjemikere lærer mer og mer om ikke-reduserbar kompleksitet og spesifiserte, komplekse nano-molekylære maskiner, nødvendige for livet blir det mindre og mindre troverdig å betrakte celler som tilfeldige biprodukt av ikke-styrte krefter. Om noe trenger å retirere i lys av vitenskapelig framgang, er det Darwinismen. Men Collins ser det ikke slik:
'Overveldende bevis'
Collins hevder at dataene gir "kraftige bevis for Darwins evolusjonsteori, som innebærer nedstamning fra en felles stamfar, ved at naturlig utvalg virker på tilfeldige variasjoner." Om mutasjoner i DNA skriver han: "Selv om de fleste mutasjoner er nøytrale eller skadelige, så vil ved sjeldne anledninger en mutasjon oppstå som vil ha en ørliten høyere sannsynlighet for å bli overlevert til framtidig avkom. Over en meget lang tidsperiode, kan slike fordelaktige sjeldne begivenheter bli spredt ut blant samtlige medlemmer av populasjonen, og til slutt føre til større endringer i biologisk funksjonalitet." (7)
Selvsagt tviler ingen på at mutasjoner kan lede til mindre endringer innen eksisterende arter (mikroevolusjon). Det beste eksempelet er antibiotika-resistens hos bakterier. Men kan tilsvarende endringer lede til nye organer, kropps-planer eller nye arter (makroevolusjon)? Allerede i 1937 kommenterte biolog Th. Dobzhansky at "Det er ingen vei mot en forståelse av mekanismene ved makroevolusjonære endringer, som krever tid på en geologisk skala, annet enn en full forståelse av mikroevolusjonære endringsprosesser, innen spennvidden til et menneskes livstid'. Han konkluderte: "Av denne grunn er vi presset .. til å sette et likhetstegn mellom mekanismene til mikro og makro-evolusjon. Så må vi fortsette ut fra denne formodningen, og pushe vår undersøkelse så lang framover som denne arbeidshypotesen tillater.
Bilde 4 Quinine-resistent
En formodning er noe, bevis noe annet. Collins skriver: "Noen kritikere av darwinismen liker å framheve at det ikke er noe bevis for makroevolusjon (større endringer i artene), bare for mikroevolusjon (stegvise endringer innen en art). Men han skriver at denne skjelningen i økende grad er kunstig." (9) For å bevise sitt poeng trekker Collins fram en studie fra Stanford angående stingsild. Stingsild i havet har typisk beskyttende plater fra hode til hale, mens i ferskvann mangler disse. Biologer har funnet en korrelasjon mellom dette og forskjellen og variasjon i genet til Ectodysplasin (EDA), et molekyl involvert i dannelse av platene. Collins konkluderer: " Det er ikke vanskelig å se hvordan forskjellen mellom ferskvanns og hav-stingsild, kan utvides til å omfatte all slags fisk. Skjelningen mellom mikro og makro-evolusjon synes dermed å være temmelig vilkårlig. Endringer som resulterer i nye arter er resultat av suksessive mindre endringer i sprangvise steg. (10)
Men stingsild i hav og ferskvann er av samme art (Gasterosteus aculeatus). Det er ikke noe bevis for at endring i genet for EDA ville, eller kunne, lede til opphav til nye arter, enda mindre befordre større anatomiske endringer som makro-evolusjon krever. Det samme kan sies om det andre eks. Collins frambringer mindre variasjoner i sykdoms-frambringende virus, bakterier og parasitter. Så Collins antar bare, i likhet med Dobzhansky at mikro-evolusjon kan ekstrapoleres (utvides) til makro-evolusjon. Til tross for 70 års genetisk forskning forblir denne ekstrapoleringen en formodning. Og skillet mellom dem er ikke mer 'kunstig' nå enn det var da.
Bilde 5 Nipigget-stingsild (ferskvann)
Angående felles avstamning skriver Collins at "studiet av multiple genomer' tillater evolusjonsbiologer å gjøre detaljerte sammenlikninger av vår DNA-sekvens med det til andre organismer. {Her kan det tilføyes at en har vært mye mer opptatt av likheter enn av forskjeller, noe som begynner å vise seg nå -oversetters tilføyelse}. I tillegg, om der er likheter beviser ikke det nødvendigvis felles stamfar, men kan peke på en felles (Intelligent) opprinnelse. For å fortsette med Collins: "Siden DNA-mutasjoner akkumulerer over tid, ville organismer med en nyere felles stamfar forventes å vise færre forskjeller enn organismer som skilte lag lengre tilbake. På genom-nivå kan en PC konstruere ett livstre basert kun på likheter i DNA-sekvenser. Denne analysen benytter ikke informasjon fra fossil-historien, eller fra anatomiske observasjoner av nåværende livsformer. Likevel er dets likhet til konklusjoner fra komparativ anatomi, både fra fossiler og fra nåværende observasjoner, slående like." Det siste er ganske enkelt en feilaktig påstand:
DNA-data leder ofte til konfliktfylte fylogenetiske kronologier. F.eks. viser Collins evolusjonære tre i "The language of God", at flyvende lemurer slekter på ekornlignende dyr, og kaniner og aper til mer fjerne grener. Men i en fylogenetisk studie publisert i 2002 (Proceedings of the National Acedemy of Sciences USA) viser at flyvende lemurer slekter mer på aper og de ekornlignende dyrene (tree shrews) mer er i slekt med kaniner. Dessuten er det ofte konflikt mellom fylogeneser basert på DNA i forhold til anatomiske baserte fylogeneser. For eks. for hvaler, som i 1960-årene ble påstått å stamme fra Mesonychider, en utdødd gruppe kjøttetende hovdyr. Men i 1990-årene viste molekylære fylogeneser større likhet til flodhest. I 2007 pekte J.G.M Thewissen og kolleger på at siden hvaler dukker opp i fossil-materialet 35 millioner år tidligere enn flodhesten, er det ikke sannsynlig at de to er nær relatert. I 2007 analyserte britiske forskere 181 molekylære og 49 morfologiske trær, og konkluderte med molekylære og morfologiske fylogeneser ofte er i strid med hverandre.
Junk-DNA?
 Collins holdt seg til tradisjonell Darwinistisk teori, som betrakter mesteparten av DNA som søppel-DNA. Da molekylærbiologer på 1970-tallet fant at menneskelig genom i all hovedsak besto av DNA som ikke kodet for proteiner, antok noen at det i hovedsak var molekylært søppel som hadde samlet seg i tidens løp. Collins kalte visse segmenter av dette 'søppel'-DNA' for ARE's (Ancient Repetitive Elements). Collins skriver at nesten halvparten av menneskelig genom består av slik genetisk vrakgods. Til sammenligning viser ENCODE-program viser at minst 80% av menneskelig genom har funksjonelle oppgaver. Det tok 20 år før den feilaktige forutsetningen om at nesten halvparten var genetisk vrakgods, ble avslørt.
Collins holdt seg til tradisjonell Darwinistisk teori, som betrakter mesteparten av DNA som søppel-DNA. Da molekylærbiologer på 1970-tallet fant at menneskelig genom i all hovedsak besto av DNA som ikke kodet for proteiner, antok noen at det i hovedsak var molekylært søppel som hadde samlet seg i tidens løp. Collins kalte visse segmenter av dette 'søppel'-DNA' for ARE's (Ancient Repetitive Elements). Collins skriver at nesten halvparten av menneskelig genom består av slik genetisk vrakgods. Til sammenligning viser ENCODE-program viser at minst 80% av menneskelig genom har funksjonelle oppgaver. Det tok 20 år før den feilaktige forutsetningen om at nesten halvparten var genetisk vrakgods, ble avslørt.
Bilde 6. Bra bok om 'søppel-DNA'
Hoved-forutsetningen for Collins var at ARE's er uten funksjon. Han skriver: "..prosessen med ombytting ødelegger ofte overførbare (hoppende) gener. Det er ARE's gjennom menneskelig og mus-genom, som ble trunkert da de slo seg ned, noe som fjernet enhver mulighet for funksjonsevne. Men nyere DNA-forskning viser at mye såkalt søppel-DNA har funksjoner. I 2006 viste japanske og amerikanske forskere at 'et stort antall ikke-kodende regioner i genomet er under sterkt selektivt press. Det innebærer at de har funksjon, eller ville ikke naturlig seleksjon påvirke dem. (13) M.a.o. det ' dysfunksjonelle' ombyttbare elementene som Collins ser som sine beste bevis, viser seg likevel ikke å være funksjonsløst likevel.
 Et lignende resultat ble rapportert i Californske vitenskapsmenn i 2007. De undersøkte 10.402 ikke-kodende elementer av det menneskelige genom og fant at en forbausende høye prosentandel fungerte i forbindelse med regulering av gener. De konkluderte med at mobile elementer kan ha spilt en større rolle enn tidligere gjenkjent. (14) det samme året kunne australske molekylær-biologer rapportere: "Mens mindre enn 1,5% av genomet koder for protein, er det nå klart at den store majoriteten blir transkribert, hovedsakelig til ikke-protein kodende RNA hvorav et stigende antall viser seg å være funksjonelle.(15) Og i 2008 demonstrerte amerikanske forskere en viktig funksjon for ikke-kodende RNA kopiert fra repeterende segmenter av DNA, som tidligere var betraktet som søppel-DNA. (16) Det synes nesten som Collins 'ufrakommelige bevis for felles avstamning' minker for hver vitenskapelig framgang.
Et lignende resultat ble rapportert i Californske vitenskapsmenn i 2007. De undersøkte 10.402 ikke-kodende elementer av det menneskelige genom og fant at en forbausende høye prosentandel fungerte i forbindelse med regulering av gener. De konkluderte med at mobile elementer kan ha spilt en større rolle enn tidligere gjenkjent. (14) det samme året kunne australske molekylær-biologer rapportere: "Mens mindre enn 1,5% av genomet koder for protein, er det nå klart at den store majoriteten blir transkribert, hovedsakelig til ikke-protein kodende RNA hvorav et stigende antall viser seg å være funksjonelle.(15) Og i 2008 demonstrerte amerikanske forskere en viktig funksjon for ikke-kodende RNA kopiert fra repeterende segmenter av DNA, som tidligere var betraktet som søppel-DNA. (16) Det synes nesten som Collins 'ufrakommelige bevis for felles avstamning' minker for hver vitenskapelig framgang.
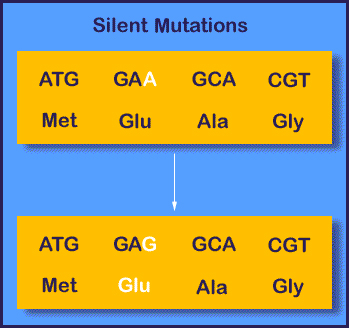
I tillegg til søppel-DNA henviste Collins også til 'stille mutasjoner'. Siden det er tre 'bokstaver' i ett kodon som spesifiserer DNA, mens totalt 4 DNA-bokstaver, innebærer det at de kan ordnes på 4!=64 måter, mens det bare er 20 aminosyrer. De fleste aminosyrer kan da bli spesifisert av mer enn én aminosyre-kombinasjon. Selv i protein-kodende sekvenser av genomet, så finnes mutasjoner som ikke endrer sekvensen av aminosyrer. Disse kalles av og til for stille mutasjoner.
I følge evolusjonsteorien skulle naturlig seleksjon i evolusjonens forløp tendere til å endre prosesser som har blitt ødelagt av endringer. Så organismer vil mer sannsynlig bære DNA mutasjoner som ikke frambringer slike ødeleggende endringer, enn av de som gjør det. I følge Collins skulle stille mutasjoner være funksjonsløse. Men en studie i 2007 av forskere ved U.S. National Cancer Institute fant at 'stille' mutasjoner i pattedyr-celler signifikant endret egenskapene ved et 'multi-drug-resistent' protein, selv om aminosyresekvensen var uendret. Om stille mutasjoner likevel har merkbar funksjon så faller Collins argument fra hverandre.
Bilde 7 Eks. på Stille mutasjon
Avrunding
 Collins hevder at han baserer sin sak for darwinismen på ny kunnskap fra genom- forskning, men han baserer seg på feilaktige forutsetninger: i) at punkt-mutasjoner og naturlig utvalg har forklart makro-evolusjon ii) at DNA-fylogeneser er overensstemmende og til å stole på og iii) at mesteparten av vår DNA er søppel-DNA. I det siste tilfellet argumenterer Collins med 'evolusjon i hullene'. Collins ord snur seg mot ham og møter ham i døra. Collins som hevdet at 'evolusjonslæren kunne og måtte være sann', møter store problemer når vitenskapen gjør framsteg. Lik mange andre ideer i fortiden, så har teistisk evolusjon knyttet vogna si til en fallende stjerne. Teistisk darwinisme eller BioLogos som Collins kaller det, passer inn i denne tradisjonen og imøtegår samme utgang.
Collins hevder at han baserer sin sak for darwinismen på ny kunnskap fra genom- forskning, men han baserer seg på feilaktige forutsetninger: i) at punkt-mutasjoner og naturlig utvalg har forklart makro-evolusjon ii) at DNA-fylogeneser er overensstemmende og til å stole på og iii) at mesteparten av vår DNA er søppel-DNA. I det siste tilfellet argumenterer Collins med 'evolusjon i hullene'. Collins ord snur seg mot ham og møter ham i døra. Collins som hevdet at 'evolusjonslæren kunne og måtte være sann', møter store problemer når vitenskapen gjør framsteg. Lik mange andre ideer i fortiden, så har teistisk evolusjon knyttet vogna si til en fallende stjerne. Teistisk darwinisme eller BioLogos som Collins kaller det, passer inn i denne tradisjonen og imøtegår samme utgang.
Bilde 8 Alternativ: Darwin in the gaps
Referanser:
1) -fra God and Evolution; J.Wells
2) https://biologos.org/
3) http://www.psykologibloggen.no/?p=2006
4) http://www.wsj.com/articles/eric-metaxas-science-increasingly-makes-the-case-for-god-1419544568
5) Behe, Darwins Black Box: The Biochemical Challenge to Evolution (New York: Free Press, 1996), s39
6) Behe, "Afterword"; Darwins Black Box: The Biochemical Challenge to Evolution (New York: Free Press, 1996), s268.
7) Collins The Language of God, s.127-131
8) Th. Dobzhansky, Genetics and the Origin og Species, preprinted 1982 (New York Columbia Un. Press, 1937), p.12
9) Collins The Language of God, s131-132
10) Collins The Language of God, s.132
11) Abstrakt tilgjengelig på: http://www.springerlink.com/content/k452146636811u14/
12) http://www.genome.gov/encode/
13) H. Hishihara, A.F.A Smit og N. Okdaa, “Functional noncoding sequences dreived from SINEs in the mammalian genome, “ Genome Research 16 (2006):s865-874,
http://www.genome.org/cgi/content/full/16/7/864
14) C.B.Lowe, G.Bejerano og D.Haussler, “Thousands of human mobile elements fragments undergo strong purifying selection near development genes,” Proceedings of the National Acedemy of Sciences USA 104(2007): s8005-8010 http://www.pnas.org/cgi/content/full/104/19/8005
15) M.Pheasant og J.S.Mattick, “Raising the estimate of functional human sequences," Genome Research 17 (2007): S.1245-1253 http://www.genome.org/cgi/content/full/17/9/1245
16)Abstract: http://www.ncbi.nlm.nih.gov/pubmed/17185560?ordinalpos=58&itool=EntrezSystem2.PEntrez.Pubmed.Pubme_ResultsPanel.PumbedRVDocSum
Oversatt og omsatt til .htm-format av Asbjørn Lund.
{kind=link}